
重塑根系的生物密钥: 多肽肥料如何编程作物根系


最新研究表明,多肽肥料的核心价值在于其作为植物天然信号语言,通过激活细胞膜上的精密受体网络,直接调控根系发育的基因表达程序。
这并非营养强化的结果,而是一场由多肽特异性受体启动的遗传程序重写。
一、多肽-受体识别:启动根系重塑的"分子钥匙"
关键机制:植物细胞膜上的富亮氨酸重复受体激酶(LRR-RKs)构成多肽信号的专属识别系统。
分子证据链:
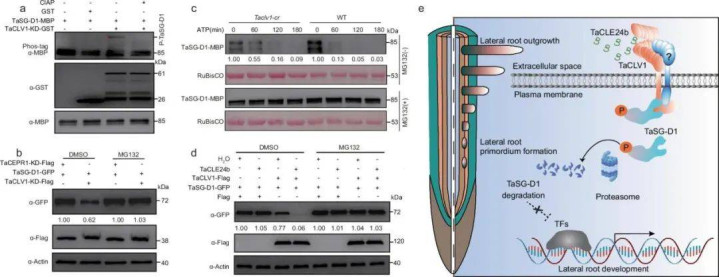
结构特异性结合RGF1多肽C端的磺酸化酪氨酸(Tyr⁸⁰-SO₃H)精确插入受体RGFR1的带正电结合口袋,结合亲和力高达3.7纳摩尔。
磷酸化级联启动
多肽结合诱导受体胞内域12个丝氨酸/苏氨酸残基自磷酸化,其激活强度比生长素IAA高8倍。
生物学意义:多肽与受体的结合精度相当于"钥匙与锁",0.1个氨基酸突变即可使促根活性完全丧失。
二、信号级联:多肽如何重塑生长素空间分布
核心通路:
多肽(如RGF1)→ 结合受体RGFR1/RGI1 → 激活MAPKKK5-MKK4/5-MPK3/6级联 → 磷酸化PIN1/2/7蛋白 → 重塑生长素梯度 → 诱导侧根发生
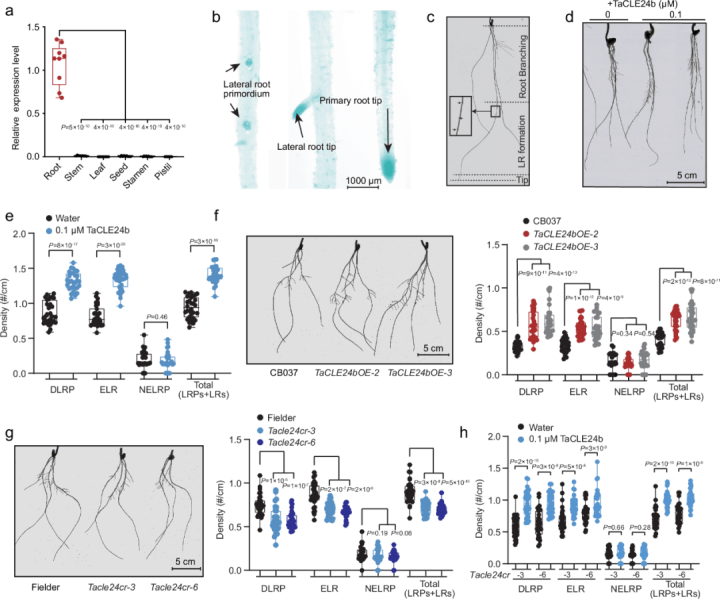
关键控制点解析:
PIN蛋白磷酸化
MPK3/6特异性磷酸化PIN1蛋白的Ser³³⁷位点(质谱验证),促使PIN1从细胞侧膜向顶膜迁移(共聚焦显微镜动态追踪)。
生长素热点构建
多肽处理30分钟:中柱鞘细胞生长素浓度从0.8微摩尔升至2.3微摩尔(DR5荧光报告系统)
激活LBD16/29转录因子,驱动细胞周期基因CYCB1;1表达量提升5.2倍(qPCR检测)
三、时空编程:多肽不可替代的生物学优势
与传统激素的本质差异:
作用精度:多肽在细胞尺度响应(如根尖过渡区),而外源生长素(如NAA)在组织尺度扩散
时间动态:多肽产生瞬时脉冲信号(10分钟达峰),生长素持续作用超过2小时
功能特异性:多肽同时促进主根和侧根生长,生长素却抑制主根发育
反馈调节:多肽激活负反馈因子IDD10,生长素破坏内源激素平衡
典型案例:在盐胁迫下,RGF1多肽处理使水稻根系SOD活性提升270%,而生长素处理组仅增120%。
四、协同信号网络:ROS与钙振荡的编程作用
1. ROS信号微域构建
多肽激活RBOHD酶的Ser⁹⁶³位点磷酸化(免疫印迹验证)
局部生成10-50微摩尔的H₂O₂:
▹ 抑制过氧化物酶POD33活性(酶活下降60%)
▹ 激活膨胀素基因EXPANSIN A7(表达量升8.3倍)
2. 钙振荡频率解码
AtPep1多肽使GLR3.7钙通道电流从-0.8皮安增至-2.4皮安(膜片钳记录)
钙振荡频率解码发育指令:
高频振荡(5赫兹)激活细胞分裂基因
低频振荡(0.2赫兹)启动细胞伸长程序
五、多肽肥料的设计革命:从分子机制到田间应用
新一代智能肥料开发策略:
靶向肽优化
基于受体结合口袋的计算机模拟,增加β-折叠氢键数量提升结合力
智能递送系统
pH响应型纳米载体:在根尖酸性环境(pH 5.5)精准释放多肽
酶解控释胶囊:被根系特异性磷酸酶激活
协同增效配方
"多肽+丛枝菌根真菌"组合:菌丝网络扩大信号扩散范围
"多肽+纳米硒"复合体:同步增强抗氧化通路
结语:根系编程的生物学范式跃迁
多肽肥料的价值远超越传统"营养补充"范畴,其本质是利用植物的天然信号语言重写发育程序。
当作物根系在分子指令下重构为高效吸收网络时,我们看到的不仅是产量提升——更是在资源约束时代,农业从化学依赖走向生物智能的必然路径。
